|
||||||
|
||||||
DOCUMENTS
elab.immagini
galileo
realtà virtuale
vrml
biomeccanica
esapodi
formula1
intelligenza
Papers
meccanica
sistemi
robotica
Come si è detto, l'intelligenza può essere definita come intuizione (insight) che prescinde da immediati riscontri esperienzali, per quanto in un organismo superiore è azzardato tentare di prescindere da ciò che è stato appreso. In ogni caso, nel comportamento manifestato dalla scimmia Sultano, certamente c'è qualcosa che rivela capacità di adattamento piuttosto fini. La capacità di adattamento che in genere manifesta ogni singolo organismo, nel tentativo di soddisfare i suoi bisogni in un ambiente vitale, viene definita normalmente in funzione delle attitudini all'apprendimento inserite nella possibilità consentite dal bagaglio genetico dell'organismo stesso. L'intelligenza sarebbe, pertanto, la risultante degli effetti prodotti dalla memoria genetica specie-specifica e di quelli indotti dall'esperienza individuale sul singolo organismo; questi ultimi vengono generalmente indicati come apprendimento.
Il fenomeno dell'apprendimento crea un importante paradosso. Sembra una forza che nega l'evoluzione! Quanto più l'apprendimento è importante nel repertorio di una specie, tanto meno sembrerebbe evolversi il comportamento di questa. Questo paradosso è stato discusso ampiamente negli scritti di Niko Tinbergen, Peter Marler, Sherwood Washburn, Hans Kummer e altri. Ciò che si evolve è il carattere orientato dell'apprendimento: la relativa facilità con cui certe osservazioni sono eseguite e certi atti sono appresi, e altri sono evitati anche in presenza di un forte rinforzo. Pavlov era in errore quando ipotizzò che qualunque fenomeno naturale scelto a piacimento può essere convertito in uno stimolo condizionato.
Se per intelligenza intendiamo genericamente delle capacità mentali strategiche quali l'astrazione dei dati concreti, la generalizzazione di alcune qualità astratte e la produzione di categorie orientative per il futuro (previsionali), per apprendimento intenderemo, più particolarmente, l'acquisizione degli elementi o contenuti concreti nuovi, anche se in maniera più o meno casuale; pur senza dimenticare che l'intelligenza e apprendimento sono due componenti fenotipiche, indicative di due varietà di un'unica modalità comportamentale. Per fare degli esempi, si può considerare il comportamento di un ratto in un labirinto: il ratto è affamato e cerca il cibo, ma non sa dov'è. Può essere a destra o a sinistra, dopo una prima o seconda o una terza svolta, ecc. Dopo un certo numero di prove, eseguite dall'animale a caso esso apprenderà il percorso adeguato è più diretto per trovare il cibo. In una gabbia di Skinner più o meno avviene lo stesso fenomeno: dopo un certo tempo l'animale apprende, piuttosto alla cieca, ciò che deve fare per raggiungere l'obiettivo (una ricompensa). Questi sono esempi di ciò che si suole chiamare apprendimento.
Nel capitolo precedente abbiamo esaminato dei casi che più propriamente possono essere indicati come intelligenza. Richiamiamo qui un esempio di Koehler, riportato in Canestrari (1984):
"In una situazione sperimentale il cibo è posto in alto e l'animale può raggiungerlo trascinandovi sotto una cassetta sulla quale montare. Mentre uno scimpanzé sta risolvendo il problema, un altro l'osserva; quest'ultimo, sottoposto a sua volta alla prova, non riesce a superarla, dato il suo basso livello intellettuale, constatato anche in altre occasioni. La sua prestazione migliore consiste nel montare sulla cassetta, ma senza averla trasportata sotto la banana."
Questa scimmia ha appreso, per osservazione e con buona imitazione, a montare sulla cassetta, senza aver però capito il perché: cioè senza avere una visione strategica del campo fenomenico totale e della sua segmentazione, per cui le è impossibile compiere una adeguata ristrutturazione del campo e quindi risolvere il problema di raggiungere il cibo.
L'apprendimento è generalmente definito come un cambiamento relativamente persistente della condotta di un animale dovuto all'esperienza. In questo l' apprendimento va distinto dallo sviluppo e dall'invecchiamento che, per quel che riguarda l'aspetti fenotipo del comportamento, sono processi analoghi ma di natura diversa. In linea generale l'apprendimento può essere inteso come in processo di informazione circa l'ambiente in cui l'animale si trova a vivere; informazioni che, integrandosi con quella contenuta nel bagaglio genetico dell'animale e in quello dell'esperienza personale e "culturale", ne rende il comportamento più adatto a quel particolare ambiente e quindi ne aumenta la possibilità di sopravvivenza. E' forse opportuno ricordare, a questo punto, che il comportamento rappresenta l'attività in toto di un organismo in risposta a stimoli interni ed esterni. Pertanto l'apprendimento, così come è stato definito, corrisponde ad un meccanismo di importanza fondamentale nell'ambito delle manifestazioni comportamentali a qualsiasi livello di organizzazione. Brevemente si può dire che il comportamento, a qualsiasi livello di organizzazione è il risultato dell'interazione di tre determinanti (Seay & Gottfried, 1978):
1. DETERMINANTE FILOGENETICO: dovuto all'eredità genetica che l'individuo riceve all'atto del suo concepimento e che è caratteristica della specie. In tal caso i membri della stessa specie condividono caratteristiche codificate geneticamente che determinano parzialmente il loro repertorio comportamentale.
2. DETERMINANTE ONTOGENETICO. influenza il livello di maturazione raggiunto dall'animale durante il suo sviluppo, per cui nel corso della vita ogni organismo modifica le sue tendenze e caratteristiche fenotipiche e quindi anche il suo comportamento.
3. DETERMINANTE
DELL'INDIVIDUO che si riferisce alle caratteristiche uniche di ciascun
organismo prodotte dalla cosiddetta variabilità genetica e dal fatto
che l'ambiente produce effetti diversi su ogni individuo, dovuti all'influenza
del passato remoto e immediato sul comportamento dell'organismo; e
ai condizionamenti culturali, per cui gli appartenenti ad un gruppo
sociale condividono un comune corpo di esperienze ambientali che tendono
a risultare in repertori comportamentali simili e in rapporti di scambio
e di informazioni.
Secondo Batacchi e Giovannelli (1988):
" dovunque c'è vita c'è crescita e sviluppo che seguono delle leggi generali, per scoprirle bisogna individuare le caratteristiche complessive dei diversi livelli evolutivi e le relazioni genetiche fra i livelli, ed estrarre delle tendenze generali dal confronto dei dati relativi allo sviluppo, individuale (ontogenesi), allo sviluppo sociale (etnogenesi), all'evoluzione della specie (filogenesi) ed alla formazione dei disturbi mentali (patogenesi)."
Esempi
di condotte comportamentali nelle quali il determinante genetico dell'apprendimento
è specifico, mentre quello acquisito individualmente dall'ambiente
è soltanto generico, si possono individuare in diversi taxa dai Molluschi
all'Homo. Per la nostra specie un esempio è costituito dal
linguaggio verbale che, attraverso le successive tappe dello sviluppo
ontogenetico, nasce da capacità specie-specifiche universali di produrre
simboli verbali in un ordine grammaticale, ereditate (filogeneticamente)
e che si concreta già nel bambino con l'acquisizione di una peculiare
lingua madre ed eventualmente di altre lingue tipiche. Ogni sociocultura
linguistica ha una sua lingua, che costituisce l'elemento ambientale
(determinante etnogenetico), il quale, tramite lo sviluppo maturazionale
del soggetto (determinante ontogenetico), va a costituire il contenuto
o uno dei contenuti della specifica facoltà (capacità o attitudine
di apprendere la produzione linguistica (determinante genetico).
Si può, quindi, distinguere fra lingua (o lingue), come sistema di dati da apprendere individualmente; e il linguaggio umano, come il sistema originale specie-specifico proprio soltanto dell'Homo sapiens sapiens, che elabora in maniera non condivisa da altre specie simboli articolati vocalmente e organizzati in strutture sematiche e in sintattico- grammaticali, solo parzialmente acquisibili da organismi filogeneticamente vicini, quali gli scimpanzé e i gorilla. Ogni uomo, alla nascita, possiede il corredo genetico adeguato per il linguaggio e, crescendo in determinati ambienti socioculturali, lo può applicare all'apprendimento di una o più lingue, mantenendo anche contemporaneamente per tutta la vita la capacità originaria di codifica universale, che gli consente anche di operare traduzioni da una singola lingua in un'altra.
Un'altra condotta specie-specifica umana, che ci consente di esemplificare il determinante ontogenetico, è costituito dal sorriso. Tale mezzo di comunicazione interpersonale non linguistico, che nell'Homo sapiens sapiens si sviluppa su base ereditaria anche i bambini sordomuti e ciechi dalla nascita (Eibl-Eibesfeldt, 1971), si modifica nei primi cinque o sei mesi di vita, in rapporto stretto con la maturazione ontogenetica da risposta istintuale a risposta socialmente acquisita e discriminante fra le categorie culturali del familiare e dell'estraneo, dell'amico e non, del simpatico e antipatico (Bilotta, Farina & Piu, 1986). Ancora un altro esempio significativo della nostra specie, è dato dalla condotta detta della babyness che potrebbe essere uno degli stimoli-segnali di un importante comportamento istintivo: l'instinto parentale (o materno), a sua volta compreso nel più vasto complesso istintuale della riproduzione della specie. La babyness sia visiva (Fullard &Reiling, 1976) che uditiva (Bertacchini, Bilotta, Farina & Piu, in stampa), è costituita da un atteggiamento emotivo affettivo e intellettivo-cognitivo aperto e positivo nei confronti dei bambini, e dei cuccioli in genere, che provoca condotte di cure parentali e comportamenti decisamente amicali. Il determinante Filogenetico di tale comportamento è molto specifico e scatta in modo appropriato solo con lo sviluppo puberale, restando, per così dire, più o meno silente per tutto l'arco dell'infanzia ; allorquando tuttavia si attiva, più prontamente nelle femmine che nei maschi, viene condizionato in maniera rilevante da determinanti individuali legati soprattutto al ruolo socio-sessuale dei soggetti, per cui poi in effetti geneticamente programmato.
Altri esempi nell'ambito dell'ordine tassonomico dei Primati ci vengono dal lavoro di Beck (1980) sull'uso di strumenti da parte degli animali. La classica ricerca di Koehler (1917) sull'uso di arnesi negli scimpanzé in cattività, è sovente citata in quanto dimostra la parte svolta dall'intuizione, opposta all'apprendimento per tentativi ed errori, nell'origine dell'uso degli arnesi. Yerkes e i suoi collaboratori hanno riscontrato anch'essi in scimpanzé in cattività alcuni casi di soluzione completa e subitanea di problemi relativi al procurarsi con un paletto del cibo posto in un tubo e all'accatastare cassette ( Bingham, 1929; Yerkes e Yerkes, 1929; Yerkes, 1943). Essi concordano nell'affermare che tali soluzioni improvvise rappresentano un processo di apprendimento del tutto simile all'intuizione di Koehler, in quanto distinto dall'apprendimento per tentativi ed errori. Pur essendo debitori di Koehler e di Yerkes per l'introduzione della provocatoria nozione di intuizione, bisogna pur tuttavia tener conto del fatto che le loro ricerche non dimostrano in modo definitivo che nell'origine dell'uso di un arnese intervenga l'intuizione. Alcuni gibboni, in una serie programmata di problemi che comportavano l'uso di un pezzo di spago, diedero soluzioni immediate molto simili a quelle che avevano portato Koehler e Yerkes a postulare il fattore intuizione. Le soluzioni immediate, tuttavia, seguivano a periodi di attività non rivolta al problema proposto, e queste attività a sua volta succedeva a tentativi persistenti, ma senza successo, di soluzione. In ogni caso i fattori determinanti dell'apprendimento, che siano più o meno condizionati dalle circostanze ambientali, mostrano che già nei Primati non umani si pongono questioni perlomeno analoghe a quelle umane. Nelle Macache giapponesi (Macaca fuscata) una giovane femmina, di nome Imo, imparò a lavare le patate prima di mangiarle, consentendosi in tal modo di gustarle meglio e di non dover sputare in continuazione sassolini, sabbia e altre impurità. Che l'acquisizione di tale scoperta sia avvenuta per intuizione o per tentativi ed errori ha poca importanza, soprattutto rispetto al fatto che le compagne di Imo impararono con successo la pratica del lavare le patate.
L'importanza
dell'apprendimento è evidente in quasi tutti i determinanti che abbiamo
indicato ed è quindi ovvio che dal punto di vista puramente biologico
esso rappresenti uno dei più "geniali" meccanismi di adattamento all'ambiente
che nel corso della storia della vita sulla Terra si è prodotto nell'ambito
dell'evoluzione biologica.
Le formulazioni dell'apprendimento, nella storia della Scienza, sono le più disparate. E' da ricordare, fra l'altro, che le due classiche Scuole di studio del comportamento, la psicologica e l'etologica, si sono a lungo osteggiate sull'origine della plasticità dei fenomeni comportamentali ritenendo, com'è noto, che i meccanismi innati e/o quelli appresi possono interferire o meno sull'intero comportamento dell'individuo.
Benché mancassero (e mancano tuttora!) dati adeguati circa il catalogo descrittivo dell'apprendimento - inteso come processo - nelle diverse specie, l'interesse per questo fenomeno si accrebbe quando la capacità di apprendere fu considerata da alcuni studiosi come criterio per dimostrare la coscienza negli animali e collegarla al tentativo di dimostrare la continuità "mentale" tra le specie, raccogliendo così lo stimolo della teoria di Darwin. Così Thorndike (lavorando con i pesci) e Yerkes (studiando le tartarughe) hanno tentato di dimostrare la capacità di apprendimento degli animali "misurando" in tal modo la loro intelligenza. Il problema della capacità di apprendere nei diversi gruppi zoologici (Fig. 2) rimane soprattutto quando, analizzando certe modificazioni comportamentali, in animali quali le planarie o i parameci, ci si imbatte in modificazioni comportamentali che è difficile riferire all'apprendimento vero e proprio.
Per quanto riguarda le teorie psicologiche sull'apprendimento bisogna ricordare che una delle prime e più note teorie è quella formulata da Torndike. Questi ritiene che il fondamento dell'apprendimento è l'associazione tra impressioni sensibili e impulsi all'azione. Perché l'associazione viene intesa come legame, connessione o connection che si rafforza o si indebolisce nel formarsi od interrompersi di abitudini, il sistema di Thorndike è noto con il nome di Connectionism, connessionismo.
Notevolmente
diverso ed in parte opposto è il pensiero dei maggiori esponenti della
teoria della Gestalt, quali Koehler e Koffka. Secondo costoro le leggi
dell'organizzazione della percezione sono applicabili anche all'apprendimento,
che viene inteso perciò come processo di organizzazione strutturale
e non di semplice associazione.
Lo sviluppo degli studi sull'apprendimento nella psicologia dell'inizio del secolo non fu seguito allo stesso modo dallo studio che il filone biologico ha fatto contemporaneamente dei processi comportamentali. E' forse da questa sostanziale differenza di metodi e di soggetti di studio che è sorta la dicotomia innato/appreso.
La questione di ciò che è innato e ciò che è appreso nel comportamento degli organismi viventi, è stata fonte di controversie accese tra gli studiosi del comportamento animale. In realtà partendo da posizioni empiriche opposte le due maggiori scuole di comportamento del nostro secolo si sono pressoché ignorate fino a che , verso gli anni '50, i rapporti tra esse divennero estremamente tesi.
Gli psicologi
comparati rimproveravano agli etologi la mancanza di controllo e rigore
sperimentale che caratterizzava gran parte della ricerca etologica,
particolarmente carente per quanto riguardava la quantificazione e
l'analisi statistica.
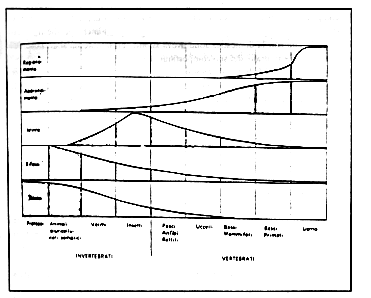
Figura 2 - Le tipologie comportamentali sono state distribuite nella scala zoologica dagli studiosi, fino ad alcuni decenni or sono, secondo un grado di complessità comportamentale crescente (da Cockrum & McCauley,1970; modificato).
Ma le critiche più feroci furono tuttavia riservate all'uso che gli etologi facevano del concetto di istinto. I primi etologi ritenevano che le basi istintive del comportamento potessero essere dimostrate con gli esperimenti di deprivazione, senza peraltro tener conto di altri fattori, interni o esterni, che possono influenzare lo sviluppo del comportamento. A tale modo di procedere reagirono con grande vigore i behavioristi e gli psicologi comparati dell'American Museum of Natural History di New York, sottolineando il fatto che l'esperienza ambientale doveva essere intesa in senso allargato e che nella produzione dei comportamenti complessi interagiscono numerosi fattori, spesso non controllati a sufficienza negli esperimenti di isolamento. D'altra parte gli etologi criticavano l'interesse quasi esclusivo degli psicologi animali per l'apprendimento dei ratti ritenendo che ciò fosse oltremodo limitante, non solo perché riguardava una sola specie, ma anche perché riguardava una specie che con la domesticazione aveva subito certamente un'alterazione delle caratteristiche "naturali" del suo comportamento.
Alcuni anni fa le scuole hanno cercato di riavvicinarsi ritenendo che le loro posizioni rigide del passato certamente non avrebbero giovato al tentativo comune di raggiungere una "sintesi" organica nello studio e nelle metodologie di approccio al comportamento soprattutto alla luce della teoria evoluzionistica. In generale oggi si ritiene che l'apprendimento costituisca un'alternativa integrata dell'istinto, che è l'informazione trasmessa geneticamente da una generazione all'altra. Anche se l'apprendimento, diversamente dall'istinto, implica di solito decisioni consce che riguardano quando e che cosa apprendere, l'autentica differenza fra l'uno e l'altro processo (e non comportamento) sta fondamentalmente nel fatto che l'istinto è un'informazione specie-specifica trasmessa geneticamente, mentre l'apprendimento è un'informazione assimilata individualmente, singolarmente, personalmente dall'ambiente esterno all'organismo. D'altra parte il lavoro compiuto negli ultimi vent'anni ha dimostrato che non può essere operata una distinzione così netta tra istinto ed apprendimento nonché tra le motivazioni che sottostanno al comportamento.
Riferendoci più in dettaglio alla posizione che la psicologia comparata ha assunto fino a pochi anni fa restano da precisare alcune cose. In generale gli studiosi di tale scuola, e anche altri - per così dire non allineati - hanno ritenuto che le differenze di apprendimento tra le specie fossero solamente di tipo quantitativo e non qualitativo. In altri termini, il processo di apprendimento era ritenuto unitario nell'ambito delle modalità di realizzazione e le differenze riscontrabili tra le specie erano dovute maggiormente alla quantità di informazione nuova e quindi appresa che una data specie era in grado di ritenere. A tale posizione si sono opposti diversi ricercatori. Si accetta, in generale, il fatto che le curve di apprendimento basate su compiti relativamente semplici sono omogenee tra le varie specie: in un semplice compito discriminativo non vi sono differenze evidenti tra la curva di apprendimento della scimmia e di un ratto. Ciò, in effetti, ha indotto a far pensare a una sostanziale continuità dell'intelligenza attraverso la filogenesi. I processi di base resterebbero appunto costanti e le differenze sarebbero dovute solo al perfezionamento dei processi primitivi e alla moltiplicazione delle strutture nervose.
Bitterman ha elaborato per le sue indagini una serie di compiti sperimentali complessi, tali da evidenziare più facilmente l'eventuale esistenza di diversi tipi di intelligenza ed il funzionamento di diversi meccanismi nervosi in soggetti di specie diverse. Esperimenti sull'inversione dell'abitudine (rinforzo della scelta per una tra due possibili alternative e successiva inversione) e sull'apprendimento di probabilità (un'alternativa è premiata l'80% delle prove e l'altra il 20%) hanno mostrato che le capacità di apprendimento sono assai diverse tra ratti e scimmie, da una parte, e pesci dall'altra. La posizione di Bitterman, che ha introdotto una nuova chiave di lettura e di metodo negli studi sperimentali sull'apprendimento, è stata criticata vigorosamente da diversi autori e la soluzione del problema è ancora abbastanza lontana.
In ogni
caso le capacità di apprendere, di utilizzare cioè le esperienze passate
per modificare il proprio comportamento in rapporto ad esse in modo
tale da raggiungere un migliore adattamento, è un requisito fondamentale
per ottenere un'elevata integrazione con l'ambiente fisico e sociale,
in continua mutazione. Se, per assurdo, il comportamento dovesse essere
geneticamente determinato in modo assolutamente rigido, esso sarebbe
indubbiamente più prevedibile, omogeneo e descrivibile con leggi rigorose,
ma sarebbe con ogni probabilità assai meno adattivo. Questa minore
idoneità biologica sarebbe un prodotto inevitabile della lentezza
con cui, di necessità, il messaggio genetico si può sviluppare legato
come è ai tempi biologici di mutazione, selezione e riproduzione:
il comportamento che ne deriverebbe sarebbe assai stereotipato, con
grandi somiglianze tra le diverse generazioni ed all'interno delle
linee parentali e sarebbe quindi ineluttabilmente assai lontano dalla
plasticità comportamentale caratteristica di gran parte delle specie
animali.
"Comunque
sia,protagonista di quasi tutti i modelli di sviluppo è l'apprendimento.
Tutti gli autori ne convengono, anche se alcuni lo denominano in modo
diverso (esperienza, sviluppo storico, sviluppo cognitivo e simili).
Il fattore apprendimento è presente in tutte quelle teorie dello sviluppo
che tengono conto di una banale prassi epistemologica scientifica
ed un ruolo diverso, spesso non preminente" (Perini, 1979).
Anche Elen Gagnè (1989) ritiene che i processi di apprendimento siano centrali nelle teorie dello sviluppo psichico, soprattutto umano: in quanto noi impariamo in continuazione dalla nascita alla morte e abbiamo una capacità di adattamento quasi illimitata.
Ciò nonostante l'autrice manifesta esplicitamente l'imbarazzo del cognitivismo rispetto al fenomeno dell' apprendimento, per lo meno a livello teoretico:
"La psicologia cognitiva dell'apprendimento è realmente agli albori della sua storia. Non deve sorprendere, pertanto, che la maggior parte dei testi di psicologia cognitiva non trattino questo argomento" (Gagné, 1989).
Noi riteniamo che possa essere un imbarazzo oggi generalizzabile ad ogni psicologo teorico, cosciente del momento di trapasso fra lo sperimentalismo, quasi sempre cieco, e la nuova sintesi che da diverse metodologie (psicologiche, etologiche, sociobiologiche, ecc.) converge sul fenomeno comportamento, illuminandone da più punti di vista le molteplici facce. Ad ogni modo c'è da sperare in un buon terzo atto della psicologia, dopo il primo atto del mentalismo illimitato e il secondo del pavlovismo e behaviorismo riduzionistici, poiché si risvegli dai sonni dogmatici.
Oggi una equilibrata visione comparatistica, ci sembra, consente di tracciare un quadro abbastanza chiaro e depurato da polemiche, fino a condurci alla formulazione di ipotesi sulla natura profonda dei fenomeni dell'apprendimento e dell'istinto, sulla probabile unificazione infine dei fenomeni comportamentali in una comune categoria fenomenlogica. Se infatti adottiamo correttamente l'ottica della comparizione fra differenti strutture organismiche e i relativi comportamenti, troviamo delle proprietà invarianti della materia vivente che come lunghissimi fili rossi compaiono in ogni forma animale ad ogni livello della scala evolutiva. Fra tali proprietà basilari dei viventi una ci interessa in particolare per il nostro discorso: si tratta della memoria sotto le due specie di memoria genetica (specie-specifica) e di memorizzazione individuale o apprendimento. Possiamo considerare la memoria specie-specifica come un programma che fornisce informazioni in larga misura standardizzate per tutti gli individui di una stessa specie, mentre l'apprendimento è un procedimento individuale che tende ad integrare l'informazione genetica con conoscenze contigenti, provenienti dall'ambiente. Lo scopo evolutivo dei due tipi di memoria è di adattare l'organismo all'ecosistema in cui vive in modo che possa crescere e riprodursi. Il crescere e la riproduzione seguono piani genetici o esigenze interne, l'adattamento è condizionato da esigenze esterne. L'apprendimento in particolare è un insieme di condotte atte ad integrare il programma genetico per evitare "incidenti" imprevisti ed imprevedibili per la memoria della specie. L'espressione comportamentale delle due memorie si modula diversamente in relazione alla differente complessità dei differenti sistemi organismici. Ma pur attraverso la grande diversità fenomenologica delle condotte fenotipiche, le due forme di memoria sembrano sostanzialmente unificabili in un unico meccanismo: la base molecolare e la relativa dinamica degli acidi nucleici, DNA e RNA.
Il DNA conserva nei geni la memoria della specie e tramite meccanismi di trascrizione diretta trasmette le informazioni che l'RNA trasferisce fuori dal nucleo e che, con la sintesi proteica, si traducono in forme fenotipiche, somatiche e comportamentali. Analogamente la memoria individuale appresa è l'informazione che dall'ambiente ecologico viene percepita, codificata e conservata forse anche negli acidi nucleici, probabilmente tramite i meccanismi di trascrizione inversa, e quindi all'occasione richiamata ed utilizzata.
Ancora Wilson (1975) scrive:
"(...) il processo di apprendimento non è un carattere fondamentale che a poco a poco insieme all'evoluzione di un encefalo di maggiori dimensioni. Piuttosto è una diversa organizzazione di peculiari adattamenti comportamentali, molti dei quali si sono evoluti ripetutamente ed indipendentemente in diversi dei principali taxa di animali".
Si può andare più in la se si pensa che i processi di apprendimento (e memorizzazione) sono proprietà già compiutamente possedute dalla catene di macromolecole organiche che chiamiamo acidi nucleici (RNA e DNA); molto prima quindi del livello cellulare. Alcuni sistemi organici, infatti, quali virus e batteri, sono perfettamente in grado, per mezzo delle loro catene molecolari di DNA e RNA, di adattarsi ad ambienti mutevoli "acquisendone" le caratteristiche, memorizzandole e modificandosi di conseguenza. Quando il processo evolutivo comincia a produrre la cellula, già da molto tempo la materia organica è dotata di strutture e di funzioni, che negli organismi superiori siamo soliti chiamare con l'etichetta generica di apprendimento-memoria. Quindi le facoltà di apprendimento di tutti gli organismi probabilmente derivano essenzialmente dalle loro dotazioni di acidi nucleici, mentre le diverse capacità di apprendere, sia quantitativamente che qualitativamente, dipendono dalle strutture o, come si dice oggi, dalle architetture delle reti neurali e qui si innesta l'argomento Wilson, appena citato, perché in realtà con il termine generico di apprendimento noi etichettiamo molti e diversi livelli di condotte fenomeniche che sembrano unificate da caratteristiche di memorizzazione a lungo termine, e, quindi, di adattamento ambientale. Aoki e Skiekevitz (1989) su tale tipo di argomenti si esprimono così:
"(...)
il programma genetico messo in atto durante la gravidanza provvede
il neonato di organi di senso pienamente sviluppati e di un cervello
ben formato, i cui 100 miliardi di neuroni sono già collegati in circuiti
per l'adempimento di funzioni specifiche. Ma prima che il cervello
in fase di sviluppo possa acquisire la piena capacità di elaborare
ed analizzare l'esperienza sensoriale, l'esperienza stessa dovrà lasciare
una sua impronta sui tessuti cerebrali".
Diversi studiosi ritengono che i processi dell'apprendimento quotidiano, anche come esperienza banale che provocano a livello fisiologico flussi di attivazione, tramite i potenziali di azione neurale, proprio in questo modo riescono a modulare il controllo del numero delle sinapsi, la quantità dell'efficienza della trasmissione intersinaptica e anche la topografia dei collegamenti sinaptici nuovi. Infatti è ormai dimostrato sperimentalmente che, se si impedisce ai neuroni neo formati di produrre potenziali di azione, le loro terminazioni assoniche si bloccano, restando immature, non produttive di nuove comunicazioni sinaptiche, né di proliferazione delle terminazioni presinaptiche. Sembra proprio che il sistema nervoso si alimenti della sua stessa attività informatica, come se l'attività creasse l'organo. Abbiamo quindi sia per l'imprinting che per l'apprendimento secondario o per gli addestramenti specialistici, un meccanismo di fondo che lavora sicuramente su almeno tre modalità: una di tipo biochimico molecolare, un'altra di tipo biolettrico di trasmissione di flussi di impulsi, una terza di tipo organizzativo delle qualità (più che quantità) del materiale mnemonico da immagazzinare in modo immodificabile (imprinting) o in maniera più o meno plastica.
La memoria a lungo termine è un processo analogo alla memoria genetica; potrebbe essere anche omologa. Cosa impedisce che gli altri processi generino DNA, anche se in forme diverse?
Il DNA è una struttura che garantisce certe proprietà alla materia vivente quali la riproduzione, la crescita, la memoria. Alla domanda di Varmus (1987): "per quale motivo la trascrizione inversa fa parte del programma funzionale di una cellula?" si può idealmente rispondere: per dotare la cellula di memoria a lungo termine!
La memoria a lungo termine è il fattore di base dell'apprendimento "psicologico" (non assuefazione, né sensibilizzazione): l'oggetto di apprendimento viene assimilato tramite memorizzazione e perciò iscritto (tramite trascrizione inversa) in qualche parte del genoma non codificante per i geni (introni; depositi di RNA?). Ecco perché in realtà non vi è salto di qualità fra ciò che si è sempre etichettato come istintuale e ciò che si considera appreso o acquisito. Istinto e apprendimento non sarebbero quindi che condotte complementari e non antitetiche, manifestazioni fenotipiche divergenti di uno stesso genotipo, a sua volta composto di elementi filogenetici, ontogenetici ed etnogenetici.
Il programma specie-specifico, proprio perché l'organismo deve operare in ambiente cui non è mai completamente adattato, viene continuamente, anche se quasi mai in maniera drammatica o "frustante", inibito e convogliato da barriere ecologiche.
Quando i coniugi Brelard introdussero il concetto di constraints on learning (1961), in clima di behaviorismo imperante almeno in U.S.A., parve opportuno limitare e delimitare il fenomeno a situazioni piuttosto eccezionali. Noi oggi riteniamo sia finalmente il caso di rianalizzare il problema a fondo così da scoprire che le constrains sono un elemento costante della fenomenologia dell'apprendimento, poiché l'apprendimento puro è un artificio di laboratorio. Il fenomeno naturale è un comportamento di un organismo programmato geneticamente, il quale per ottenere soddisfazione ai suoi bisogni e raggiungere i propri scopi di sopravvivenza e riproduzione, si adatta modulando le sue condotte sulle situazioni ambientali contigenti. Anche in laboratorio non si riuscirebbe a condizionare minimamente un qualsiasi organismo, se non si sfruttassero le sue pulsioni profonde, le sue motivazioni interne e, in definitiva, l'attivismo spontaneo delle sue condotte comportamentali specie-specifiche. Basta, ad esempio, considerare un ratto nella gabbia di Skinner: se l'animale non fosse estraneamente attivo nell'esplorazione non scoprirebbe mai la leva o un altro apparato, che poi serve a condizionarlo. La fenomenologia della costreins on learning ci indica l'esistenza di canalizzazioni comportamentali dell'apprendimento secondo categorie di condotte filogeneticamente antiche: un ratto per condotte alimentari (mangiare e bere) impara abbastanza facilmente a premere una leva con una zampa, ma per evitare una scossa elettrica (condotta di difesa) non impara a premere una leva, bensì a saltare e correre per scappare dal pericolo. I piccioni impararono piuttosto facilmente condizionamenti che comportano il beccare, se la ricompensa è il cibo; mentre non imparano gran ché se devono beccare per evitare una scossa elettrica. L'evitamento della scossa elettrica può essere appreso bene dai piccioni se la risposta condizionata prevede il saltare su e giù con le zampe; al contrario tale condotta non aiuta ad apprendere le soluzioni di un compito da alimentazione. Le api da miele imparano a riconoscere i fiori da cui trarre bottini di nettare e polline, acquisendo e memorizzando delle caratteristiche legate a forme, colori e odori dei fiori stessi e trascurando completamente segnali che pure percepiscono in contesti non alimentari.
Nell'ambito dell'apprendimento operano, insomma, delle selezioni di stimoli-segnali del tutto analoghe a quelle operate in genere da tutti gli organismi nell'ambito delle condotte istintituali. Sembra che una delle caratteristiche della memoria sia di essere preorientata dalla struttura organismica che la esercita.
Gould
e Marler (1987) così si esprimono:
"(...) il lavoro compiuto negli ultimi vent'anni ha dimostrato che non può essere operata una distinzione così netta tra istinto e apprendimento nonché tra le motivazioni che sottostanno al comportamento umano ed animale. Si è riscontrato, per esempio, che molti insetti apprendono in modo prodigioso. Tuttavia ora sappiamo che il processo di apprendimento negli animali superiori, così come negli insetti, avviene spesso in modo innato, ossia regolato dall'informazione contenuta nel patrimonio genetico dell'animale. In altre parole lo stesso processo di apprendimento è spesso controllato dall'istinto".
Già nel 1974 Canestrari e Bertacchini avvertivano che "se si usa il concetto di apprendimento come contrapposto al concetto di istinto o come complemento dell'innato si dicotomizza il fenomeno comportamentale". Se si guarda il comportamento solo dall'angusto spiraglio degli artifici di laboratorio si può anche scambiare il complesso sistema organismico per una tabula rasa. In effetti la maggior parte dei fenomeno di apprendimento necessitano almeno di alcune tabulae rasae su cui iscrivere i propri effetti a lungo termine. Ma certo il sistema nervoso non è solo questo: esso possiede anche altri meccanismi e processi di adattamento ecologico. I riflessi, gli istinti, le varie forme di constrains sono strutture di adattamento quantitativamente più sviluppate, filogeneticamente antiche e quindi atte a modulare, integrare e dirigere i processi di apprendimento versi obiettivi evoluzionistici che hanno modellato lo stesso organismo. E' lecito pensare quindi che le tabulae rasae non siano solo espressioni metaforiche, ma strutture biochimiche di registrazione mnemonica molecolare, analoghe se non omologhe agli acidi nucleici che conservano la memoria genetica e decodificandola producono quei comportamenti specie-specifici che modulano appunto l'apprendimento individuale. Non vi sarebbe nulla di strano se si scoprisse che la memoria a lungo termine, come il suo omologo, la memoria genetica, venisse codificata per mezzo della trascrizione inversa nel DNA: in quelle porzioni del DNA, dette introni, che non codificano i geni. Ma questo costituirebbe solo la base (la banca dati); tutte le qualità dinamiche dell'apprendimento, che conducono al pensiero, al linguaggio, alla creatività, probabilmente dipendono "dall'architettura sia dell'hardware neurale sia dal software informatico" che viene elaborato dalle reti neuronali (Gelernter. 1989).
Nel tentativo di costruire macchine sempre più dotate di funzioni almeno analoghe a quelle del cervello biologico, si vanno scoprendo relazioni intrinseche fra le memorie di calcolatori in parallelo: i flussi di dati di apprendimento si correlano fra loro in rapporto all'architettura del sistema di calcolo. Ad alcuni ricercatori non sembra necessario impegnarsi a costruire una "mente", ma piuttosto creare dei corpi (es. robot) che automaticamente nel loro rapporto di adattamento all'ambiente, addestramento, apprendimento vadano ad organizzare livelli superiori di capacità. Clark (1987) sostiene che:
"(....) vi è un esteso convincimento che la mente possa essere studiata isolandola dai dettagli delle sue incarnazioni fisiche e dalle condizioni ambientali di questi. Questa è una forma di Cartesianismo residuale in cui la scienza cognitiva può rischiare di incorrere".
Inoltre Clark presenta una serie di argomentazioni che mostrano come i dati mentali non dovrebbero essere considerati isolati dalle capacità motorie e dalla manipolazione di oggetti che compiono le strutture somatiche. l'autore mette anche in evidenza che il rapporto soma-ambiente va visto nel suo duplice aspetto, come nella realtà ,fenomenica della biologia, di relazioni con oggetti fisici e di interrelazioni con soggetti sociali, richiamando l'attenzione sulla concretezza dei processi evolutivi naturali, che hanno condotto l'automa organismo da livelli elementari di risposte all'ambiente fino a livelli complessi o mentali.
Smolensky (1987) ci sembra sulla stessa linea teorica, quando espone la sua visione connessionistica, che tende più a risolvere i problemi pratici e a produrre modelli applicativi concreti che a descrivere i fenomeni cognitivi. Egli afferma:
"presentare una visione del connessionismo che implica che il livello di analisi a cui uniformare processi formali di conoscenza può essere individuato nel livello subsimbolico,intermedio fra i livelli neurali e quelli simbolici".
A tale livello egli pone:
"
due tipi di variabili: un livello di attivazione per ciascuna delle
unità e una forza connettiva per ciascuno dei legami (....).Le regole
di attivazione sono regole di inferenza: non di inferenza logica,
ma di inferenza statistica. Le regole di modificazione della forza
connettiva sono immagazinamenti di memoria e procedure di apprendimento(...).
La conoscenza sarebbe il risultato di sistemi dinamici e non di calcoli
digitali. Questo è il risultato di un formalismo ispirato dalla struttura
neurale piuttosto che da quella mentale (....). La categorizzazione
non è l'esecuzione di un algoritmo simbolico, ma la continua evoluzione
del sistema dinamico, l'evoluzione che pilota gli stati nelle zone
di attrazione, che soddisfano la massima armonia. L'apprendimento
non è la costruzione e l'attuazione di formule, ma l'aggiustamento
graduale di forze connettive con l'esperienza, con l'effetto di configurazioni
che lentamente raggiungono un'armonia, adattando vecchi concetti,
categorie e schemi e creandone di nuovi (...). L'unità sottostante
la conoscenza la si trova non a livello simbolico, ma piuttosto a
quello subsimbolico, dove pochi principi di un singolo quadro formale
conducono a produrre una ricca varietà di comportamenti globali"
Tale concezione dei processi di apprendimento è oggi piuttosto comune nella descrizione della psicologia fenomenica,
che spesso riassume tutto questo insieme di processi cognitivi con l'espressione "struttura cognitiva". Tale struttura è caratterizzata in maniera eminente dal suo modo dinamico di variare continuamente in rapporto alle esperienze che ogni individuo compie nella sua esistenza, sovrapponendosi quindi quasi completamente al determinante individuale dell'apprendimento e parzialmente anche al determinante ontogenetico. Ogni nuovo contenuto di apprendimento non va a stratificarsi su una tabula rasa, bensì si aggiunge ad un complesso amalgama precedente, ove raggiunge una posizione armonica. Per quello che ci dice la psicologia contemporanea risulta della massima importanza, sia per l'automa biologico che per quello artificiale, il rapporto quantità/qualità del materiale da apprendere. Rispetto alla pura quantità, se mai fosse possibile avere contenuti semplici differenziati solo per la lunghezza e numero, il fattore tempo a disposizione per apprendere sarebbe sufficiente: in tempi più lunghi si apprendono più contenuti. Ma la quantità quasi sempre (anche nelle più favorevoli condizioni quali sono quelle scolastiche) viene complicata dalla difficoltà intrinseca a certi contenuti, per cui l'assimilazione e la ritenzione possono anche non verificarsi. Quindi si può rendere necessario semplificare i contenuti complicati, in modo da dotare l'automa di nuove cognizioni gradualmente, quasi che i contenuti richiedano di "appoggiarsi e sostenersi" l'un l'altro come le sbarre di un traliccio. Perciò bisogna mettere l'automa nelle condizioni più favorevoli ad accettare certi contenuti: creargli delle motivazioni favorevoli ad apprendere ciò che si desidera apprenda. Le sue attitudini di base aumenteranno proporzionalmente all'implementazione che ne riceveranno, creando nuove capacità, anche di alone, per assimilare contenuti più complessi dei precedenti e diversi, ma adiacenti o analoghi. Raggiunte alcune attitudini minimali di apprendimento autonomo, è opportuno lasciare all'automa la possibilità di organizzare i successivi apprendimenti nel modo più autonomo possibile: i soggetti che organizzano il proprio materiale di studio con pochi suggerimenti esterni, dimostrano maggiore ritenzione e quindi autoimplementazione poiché i contenuti nuovi diventano anche nuovi mezzi strumentali di crescita cognitiva; altrimenti il materiale nuovo di apprendimento viene immagazzinato a livelli marginali, non strutturati col resto dell'acquisito. Nel primo caso si parla di ritenzione significativa, che si riferisce all'acquisizione di nozioni che aumentano la capacità di conoscenza, poiché venga a riunirsi in maniera duratura alla matrice cognitiva ove, interagendo dinamicamente con i contenuti precedenti, evolvono le capacità complessive del sistema. Nel secondo caso si dice che la ritenzione è meccanica, in quanto è memorizzata in maniera segregata e non organica col resto delle conoscenze. La superiorità cognitiva del processo di ritenzione significativa, rispetto a quella meccanica, riflette una reale differenza nella natura originaria dei rispettivi processi di memorizzazione, anche se entrambi avvengono con modalità automatiche.

