|
||||||
|
||||||
DOCUMENTS
elab.immagini
galileo
realtà virtuale
vrml
biomeccanica
esapodi
formula1
intelligenza
Papers
meccanica
sistemi
robotica
INTELLIGENZA BIOLOGICA E INTELLIGENZA ARTIFICIALE
Apprendimento ed
intelligenza in Octopus Vulgaris
L'utilizzo del polpo in situazioni sperimentali di laboratorio ha una tradizione abbastanza consolidata, sebbene gli studi fino ad oggi condotti si riferiscano soprattutto agli aspetti biologici e fisiologici di questo organismo. La capacità in Octopus di apprendimento pavloviano è ampiamente documentata in letteratura. La tecnica del condizionamento classico, con l'uso di rinforzi positivi e negativi, è stata largamente applicata da neurobiologi e fisiologi allo studio dei sistemi meccano tattile, chemiotattile e visivo del polpo, mettendone in risalto l'estremo grado di finezza dei meccanismi percettivi.
I contributi maggiori in questo campo risultano dal lavoro di M.J.Wells e di J.Z. Young. Il sistema nervoso del polpo è congegnato in modo che l'animale attacca quando un oggetto qualsiasi si muove all'interno del suo campo visivo. Ovviamente, in mancanza di ricompensa (stimolo incondizionato), si andrà incontro ad un processo di estinzione (Young, 1959), per cui la percentuale di attacchi in un certo tempo ad un dato oggetto andrà decadendo, mentre un nuovo oggetto verrà ancora attaccato. La somministrazione della ricompensa determina quello che comunemente descritto come apprendimento ad attaccare: l'effetto immediato del cibo è di accrescere la tendenza ad attaccare. Analogamente è possibile determinare un apprendimento a non attaccare un dato oggetto, somministrando una leggera scossa elettrica attraverso una coppia di elettrodi posti su di una bacchetta (Maldonado, 1965). In tal modo, il polpo può imparare a reagire opportunamente ad un'ampia gamma di situazioni visive o tattili (Mackintosh, 1965).
Il sistema visivo del polpo è uno dei più efficienti tra le specie marine. I lavori di J.Z.Young (1956,1961) e di N.S. Sutherland (1957,1958,1960,1963) sono eloquenti in proposito: i polpi imparano rapidamente ad attaccare o a non attaccare sagome diverse tra loro o diversamente orientate (ad esempio rettangoli verticali piuttosto che orizzontali e viceversa). E' stato notato che se da un lato l'Octopus distingue facilmente i rettangoli verticali da quelli orizzontali ed i triangoli dai quadrati e dai cerchi, maggiori difficoltà si dimostrano quando si tratta di distinguere i quadrati dai rombi e dai cerchi. Inoltre, gli animali sono incapaci di distinguere dei rettangoli mostrati in posizioni oblique. Il modello degli " analizzatori di caratteristiche" di Sutherland (1959), rispondendo al problema teorico di descrivere il processo cognitivo di riconoscimento, prevede che le configurazioni vengano identificate attraverso opportuni analizzatori in virtù dei loro " attributi" ( ad es. linearità, angolarità, concavità, orizzontalità, ecc.). Sulla base dei dati sperimentali, Sutherland (1961,1963) avanza l'ipotesi che il sistema analizzatore del polpo discrimina le forme unicamente in base alle dimensioni orizzontale e verticale mancando invece di analizzatori dell'obliquità: il polpo avrebbe un maggior numero di cellule sensibili alla dimensione verticale rispetto a quelle dell'orizzontale, ed entrambi questi tipi di cellule sarebbero presenti in numero maggiore di quelle per l'obliquità.
Le discriminazioni
apprese permangono diversi giorni. Young (1960, 1964) osserva come
non sia errato supporre che le connessioni neurali, una volta stabilite,
permangono molto a lungo e che il loro decadimento nella memoria sia
dovuto piuttosto a fenomeni di interferenza con altre connessioni
che determinerebbe l'impossibilità di leggere opportunamente nella
memoria (cfr. anche Sanders & Barlow, 1971).
I lavori di Wells (1960, 1964 b) e di Wells & Wells (1956) indicano ineluttabilmente come negli otto tentacoli e nelle ventose del polpo sia presente un sistema tattile meglio sviluppato di quello di qualsiasi altro invertebrato. I ricettori si trovano soprattutto sui bordi delle ventose e mandano fibre nelle catene dei gangli dei tentacoli. I gangli dei tentacoli, uno per ciascuna ventosa, sono capaci di un notevole controllo indipendente. Wells ha dimostrato la sottile capacità di discriminazione tattile usando polpi accecati mediante rescissione dei nervi ottici. La tecnica è quella nota del condizionamento pavloviano: in presenza di una ricompensa l'animale attirerà l'oggetto alla bocca, respingendolo, invece, in assenza di ricompensa o in presenza di un rinforzo negativo. E' dunque risultato evidente che i ricettori dei tentacoli sono in grado di mandare al cervello dei segnali sul grado di ruvidezza degli oggetti toccati e sulla loro natura chimica. Una elevata sensibilità chemiotattile è stata rilevata usando oggetti imbevuti in concentrazioni minime di acido cloridrico, saccarosio e chinino (Chase & Wells, 1986). Dagli esperimenti di Wells (1961 b) sulla discriminazione di peso, risulta che gli Octopus non sono in grado di apprendere ad afferrare un cilindro in plastica e a non afferrare un altro appesantito con del piombo.
Nell'ambito
dell'apprendimento a non attaccare oggetti (con la tecnica delle scosse)
è stato notato un fatto curioso: presentando, durante l'addestramento,
l'oggetto sempre allo stesso tentacolo e quindi, successivamente,
ad un altro tentacolo dalla parte opposta del corpo, esso viene inizialmente
accettato; me se le prove sono distanziate ad intervalli di mezz'ora,
l'apprendimento ha tempo di "diffondersi" a quelle parti dei centri
connesse con tutti gli altri tentacoli e l'oggetto viene respinto
da qualunque di essi.
Come
già accennato, gli studi sopra citati erano finalizzati prevalentemente
alla comprensione della neurofisiologia del polpo: Boycott & Young
(1950, 1955); Wells (1959,1961a,1966,1967,1978) e Young (1964,1965,1983),
in particolare, con la tecnica del condizionamento e mediante lesioni
selettive indotte sui soggetti sperimentali, hanno potuto individuare
alcuni centri nel sistema nervoso di Octopus che sembrerebbero
essere implicati nei diversi tipi di apprendimento (cfr. anche Wells
& Wells, 1957). In base alla mappatura che ne risulta, hanno creato
dei modelli tendenti a spiegare l'organizzazione "funzionale" del
cervello. E' interessante notare come nell'Octopus, secondo
tali modelli, l'integrazione tra sistema propriocettivo e sistema
visivo sembri essere molto ridotta (Wells, 1960), dal momento che
non sono state evidenziete connessioni tra i centri relativi ai due
sistemi di memoria, a livello di Sistema Nervoso Centrale.
Un altro aspetto delle capacità di apprendimento in Octopus riguarda quelle definite come proprietà cognitive. I primi esperimenti risalgono all'inizio del secolo: Piéron (1911) e Bierens de Haan (1926) analizzarono le risposte di Octopus ad un labirinto molto semplice, costituito da un cilindro di vetro contenente un granchio vivo. Nonostante l'estrema semplicità del compito di aggiramento richiesto, le conclusioni raggiunte dai due autori si contraddicono. Secondo Piéron il polpo impara ad aggirare l'ostacolo ed a nutrirsi infilando un tentacolo nell'imboccatura. Tuttavia sebbene l'autore si aspettasse nel corso dell'allenamento una diminuzione dei tempi di risoluzione del problema, una riduzione progressiva ed una maggiore precisione dei movimenti esplorativi, il verificarsi di questi eventi non risulta chiaramente dal suo lavoro; non si evidenziò dunque una vera performance di apprendimento e, anche se certamente si verificarono dei cambiamenti comportamentali, la risoluzione del problema avrebbe potuto avvenire a caso. Le conclusioni dell'esperimento di Bierens da Haan (1926), viceversa, affermano perentoriamente l'incapacità del polpo a risolvere il compito. Nel suo studio venne utilizzato un solo animale, il quale, dopo numerosi tentativi falliti, rinunciò definitivamente ad ogni successivo tentativo: ne risulta, dunque, un buon esempio di processo di estinzione degli attacchi. Diversa è la situazione sperimentale utilizzata da Boycott (1953) il compito richiesto era di aggirare due pareti opache seguendo un percorso a forma di Z per raggiungere il cibo. Dopo un preallenamento in cui veniva mostrato il percorso trascinando lungo di esso un granchio legato ad un filo, su 5 animali testati, solo uno di essi fu in grado di risolvere il labirinto. Nello stesso lavoro, l'autore semplificò il compito predisponendo un labirinto costituito da una sola parete da aggirare, questa volta trasparente, dietro la quale si trovava il granchio: su tre animali testati, solo uno imparò a risolvere stabilmente il compito di aggiramento, continuando tuttavia in tutte le prove (44 in totale) a tentare la via diretta, spingendo ed esplorando il vetro per una durata di tempo compresa tra 10/20 secondi. Boycott concluse che gli Octopus sono capaci di apprendere il labirinto ma che esiste una forte variabilità interindividuale.
Mentre
negli esperimenti sopra elencati il labirinto richiedeva un semplice
aggiramento di ostacoli, negli esperimenti di Schiller (1949) e Wells
(1964 a,c; 1967) il labirinto comportava una scelta multipla tra due
percorsi, due soluzioni possibili di cui una sola veniva ricompensata.
Si trattava di un labirinto a T semplificato dall'accorgimento che
il polpo era in grado di vedere dallo start-point, attraverso
un vetro, in quale dei due lati si trovava la preda. Per raggiungerla
il polpo doveva percorrere un corridoio tra due pareti opache, al
termine del quale doveva scegliere se svoltare a destra o a sinistra.
Entrambi gli autori, testando solo pochi animali, conclusero che gli
Octopus sono capaci di apprendere questo tipo di labirinto.
Circa le modalità attraverso cui tale apprendimento si realizza, Schiller
affermò che gli animali per orientarsi si baserebbero essenzialmente
su indici tattili, mantenendo costantemente il contatto con la parete
del corridoio dal lato giusto, fino ad aggiramento completato. Wells
dissentì da tale conclusione, sottolineando l'importanza degli indici
connessi all'orientamento visivo.
Sempre
utilizzando un numero limitato di Octopus, un altro gruppo
di lavori (Dews, 1959; Crancher et al., 1972) hanno avuto come oggetto
di studio l'apprendimento ad utilizzare un distributore automatico
di cibo. In Dews (1959) l'operandum era costituito da una leva che
bisognava tirare per ottenere una ricompensa: su tre animali testati,
due apprendevano il compito in maniera evidente. In Crancher et al.
(1972) il compito era costituito da un tubo a fior di acqua entro
cui l'animale doveva infilare il tentacolo per interrompere un circuito
fotoelettrico ricevendone la ricompensa. 6 Octopus furono testati
in questo lavoro anche con programmi di rinforzo a rapporto variabile
(VR), mostrando solo nel caso di un esemplare un aumento consistentemente
maggiore nel caso di VR rispetto al programma di rinforzo continuo.
Questi esperimenti dimostrano in maniera chiara la possibilità di
condizionamento operante in Octopus. La difficoltà maggiore
consisteva comunque nell'ottenere delle unità discrete di risposta:
in Dews (1959) gli animali tendenzialmente restavano aggrappati alla
leva, manifestastando risposte consumatore nei suoi confronti. Nell'esperimento
di Crancher et al. (1972) gli animali tendevano a mantenere
il tentacolo all'interno del tubo, impedendo così il successivo funzionamento
del dispositivo di distribuzione del cibo.
Data
l'esiguità e la contraddittorietà dei dati presenti in letteratura
sulle capacità di apprendimento di in Octopus in compito di
tipo strumentale ed in situazioni di apprendimento cognitivo, sono
stati realizzati presso la Stazione Zoologica di Napoli una serie
di esperimenti atti a fare luce su tale argomento.
Dopo
un periodo di digiuno i polpi sono stati nutriti utilizzando un problem-box
costituito da un barattolo in vetro chiuso da un tappo di plastica
ad incastro. La principale ipotesi era la verifica di un miglioramento
progressivo delle curve dei tempi di risoluzione del compito, tale
poter concludere che sussiste un apprendimento, e non l'eventuale
risoluzione avviene casualmente (cfr.Pieron, 1911). Inoltre, mediante
un opportuno controllo della situazione sperimentale, delle variabili
intervenienti e dei parametri chimico-fisici di base, così come sono
stati evidenziati dalle esperienze di Losito et. al. (1985),
si era interessati a verificare il grado di prevedibiltà di tale apprendimento,
l'omogeneità interindividuale delle prestazioni e l'eventualità di
costruire delle curve standard, nella più ampia prospettiva di poter
disporre di uno strumento utilizzabile in ulteriori esperimenti. Infine,
oltre ad un'analisi statistica dei risultati, già in fase di osservazione
si è disposto, grazie all'uso di videoregistrazioni, di materiale
sufficiente per eseguire un'analisi preliminare delle strategie adottate
dagli Octopus nel risolvere il compito.
Tutto
il nostro lavoro è stato condotto con l'Octopus vulgaris Cuvier:
questa è infatti una specie di Octopus, tra le settantacinque
esistenti al mondo, più comune nel Golfo di Napoli. Una volta pescati
dal Servizio Pesca della Stazione Zoologica di Napoli, gli animali
di entrambi i sessi sono stati pesati e posti in vasche speciali separate
dando a ciascuno di essi un numero. Per ciascun animale è stato stabilito
un periodo minimo di adattamento alla nuova situazione di una settimana.
Nei primi due giorni, inoltre, i polpi sono stati regolarmente nutriti,
quindi sono stati tenuti a digiuno nei seguenti 5 giorni o comunque
nei 5 giorni precedenti l'inizio dell'esperimento. Sono stati scartati
quegli animali che presentavano delle lesioni ed alcuni di essi sono
deceduti durante il periodo di adattamento o ad esperimento inoltrato.
Le vasche sperimentali (Figura 3) erano costituite da vasche di stabulazione delle dimensioni di 60X100X50 cm, con acqua di mare corrente (volume di circa 0.34 1/10 sec), ed una capacità di 250 1. Ciascuna vasca era costituita in materiale sintetico sulla sui facciata anteriore era stata ricavata una finestra di vetro (unidirezionale) 45X35 cm, per l'osservazione e le riprese.
Superiormente la vasca era chiusa da un coperchio in plexiglas in modo da consentire il passaggio della luce. L'illuminazione era costituita da lampade fluorescenti a luce bianca, in modo da mantenere un'intensità luminosa pari a quella riscontrabile a 2-5 metri di profondità. Le serie di lampade erano controllate da timer programmati in modo che il ciclo di illuminazione era in accordo a quello nictemerale, determinato sulla base delle effemeridi nautiche. All'interno della vasca trovava posto una tana costituita da mattoni.
Quotidianamente
venivano rilevati e controllati i parametri fisico-chimici dell'acqua
di mare indicati in tabella 2.
Tabella2 - Parametri chimico-fisici rilevati e controllati durante tutti gli esperimenti di apprendimento.
| Parametro | Metodo |
|
temperatura densità tasso di ossigeno tasso di ammoniaca |
termometro densimetro a gravità O2 Hilena test NH4 Sera test |
L'andamento stagionale dei suddetti parametri è stato confrontato, per tutta la durata degli esperimenti, con quello riscontrato nel Golfo di Napoli nel corso delle normali attività di ricerca del Laboratorio di Oceanografia Biologica della Stazione Zoologica di Napoli.
Il
problem-box era costituito da un cilindro in vetro dell'altezza di
14 cm, del diametro alla base di 8 cm, e del diametro all'imboccatura
di 6 cm. Il tappo era in plastica colore rosso del diametro di 5.5
cm e dello spessore di 1 cm. Il peso vuoto del barattolo era 281 g,
quello del tappo 5g. Tra una prova e l'altra veniva effettuato un
accurato lavaggio e sterilizzazione sia del cilindro che del tappo.
All'interno del barattolo veniva introdotto un granchio vivo (Carcinus
mediterraneus) del peso di circa 4 g, ed il problem-box veniva
a sua volta introdotto nella vasca sperimentale in posizione verticale.
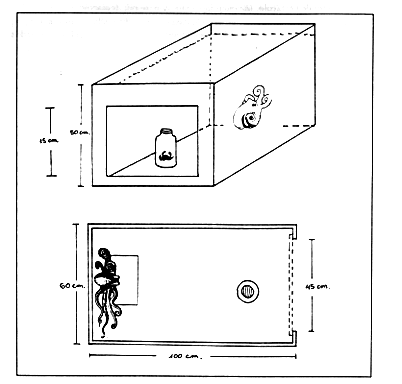
Figura
3 -Visione, in prospettiva e dall'alto, della vasca utilizzata
per l'esperimento di problem-box indicante le posizioni relative
dell'animale e del barattolo all'inizio di ogni prova.
La soluzione richiesta dal compito può essere considerata nei suoi due momenti fondamentali: a) la rimozione del tappo (azione sull'operandum) attraverso l'utilizzo delle ventose tentacolari; il movimento richiesto era ben preciso data l'impossibilità di girare il tappo operando una pressione verso il bordo; b) aggiramento dell'ostacolo (detour) infilando uno o più tentacoli all'interno del problem-box.
Di conseguenza, venivano rilevati i seguenti tempi:
Latenza di attacco: dal momento in cui il barattolo è immerso nella vasca, al momenti in cui inizia l'attacco. Nel caso di più attacchi all'interno di una stessa prova, è stata considerata la media delle latenze dei vari attacchi;
Operandum: una volta attaccato, dal momento in cui l'animale tocca il barattolo al momento in cui rimuove il tappo;
Detour: dalla rimozione del tappo, al momento in cui tocca la preda.
L'esperimento
si è svolto in tre fasi distinte. Una prima fase esperimentale si
è svolta a cavallo tra Gennaio e Febbraio 1988: 7 Octopus (anim.
n. 8, 9, 10, 11, 12, 13 e 14) di entrambi i sessi sono stati nutriti
due volte al giorno per cui la durata di 10 giorni utilizzanti il
problem-box. Le sessioni sperimentali avvenivano tra le 11:00 e le
12.00 per la mattina e tra le 16:00 e le 17:00 per il pomeriggio (cfr.
Bradley & Young, 1975). In ogni sessione venivano concesse due
opportunità a ciascun animale: il barattolo veniva lasciato nella
vasca per un tempo massimo di 5 minuti (305 sec). Se l'animale non
risolveva entro il tempo limite, veniva ritirato il barattolo per
poi essere successivamente introdotto una seconda volta dopo un tempo
di circa 15-20 minuti. i parametri chimoco-fisici registrati in questa
fase 1 sono riportati in tabella 3. Tali valori rientravano nei limiti
accettabili e soprattutto sono rimasti costanti nei 10 giorni.
Tabella 3 - Valori dei parametri chimico-fisici registrati durante la prima fase sperimentale.
|
temperatura dell'acqua 15.5° - 17° densità dell'acqua 1.023 - 1.024 tasso di NH4 < 0.02 mg/lt tasso di O2 9.5 mg/lt |
Solo la temperatura dell'acqua, come si discuterà in seguito, è da ritenersi eccessivamente bassa: il metabolismo è rallentato e , considerando anche l'inerzia comportamentale indotta dalla condizione di cattività, le necessità di cibo sono ridotte al minimo.
Nella
seconda fase sperimentale, nel mese di Aprile 1988 (tabella 4), proprio
a causa della caduta motivazionale riscontrata durante la fase 1,
si è abbassato da due ad uno il numero delle sessioni giornaliere
e quindi l'intera durata dell'esperimento è stata raddoppiata a 20
giorni. La prova si svolgeva tra le 16:00 e le 17:00 e venivano concesse
sempre due opportunità con un tempo limite di 5 minuti. Sono stati
utilizzati 5 Octopus (Anim. n. 16, 18, 21, 22 e 23), tutti
nuovi.
Tabella 4 - Valori dei parametri chimico-fisici registrati durante la seconda fase sperimentale.
|
temperatura dell'acqua 17°-19° densità dell'acqua 1.024 tasso di NH4 < 0.02 mg/lt tasso di O2 9 mg/lt |
Nella terza fase, nel mese di settembre 1988 (Tabella 5), sono stati testati 8 Octopus (Anim. n.29, 30,32,34,35,36,37e38) adottando il medesimo schema della fase 1 (due volte al giorno per 10 giorni), essendo la temperatura dell'acqua notevolmente più alta.
Per tutto il campione (n=20 animali) sono state calcolate medie, errori standard e limiti fiduciali dei tempi di Latenza di Attacco, Operandum e Detour. Sono stati anche calcolati i valori del coefficiente di correlazione r di Bravais-Pearson e le rette di regressione. E' stato, inoltre, utilizzato un programma di analisi statistica, denominato STATA, che utilizza test statistici parametrici e non parametrici, e che - tra l'altro - contiene il test di Analisi della Varianza per Trend a due vie senza repliche di Edwards (1967). I grafici e i valori tabulati sono stati ottenuti per mezzo del programma EXCEL (Microsoft, USA). I dati relativi a ciascun animale sono stati, altresì, conservati in un archivio per la gestione dei dati (Database Relazionale, RBASE, Microrim, USA) disegnato ad hoc.
Le figure
4,5 e 6 mostrano l'andamento delle medie per i tempi di apertura (Operandum)
e di cattura del granchio (Detour) calcolate sulla base delle prestazioni
dei 20 animali dei tre gruppi sperimentali osservati nel corso delle
20 prove.
Tabella 5 - Valori dei parametri chimico fisici registrati durante la terza fase sperimentale.
|
temperatura dell'acqua 24° 25° densità dell'acqua 1.026 tasso di NH4 < 0.02 mg/lt tasso di O2 8.5 mg/lt |
I dati
relativi al numero di attacchi errati e di attacchi totali indicano
che c'è un significativo decremento di attacchi errati nel corso delle
prove. Questa indicazione integra quanto risulta dall'analisi degli
andamenti dei valori medi dei tempi di Operandum e Detour (Figure,
4-6). Le prove irrisolte sono state conteggiate sulla base dei tempi
di Operandum; infatti nel caso di mancata risoluzione nel tempo massimo
consentito (5 minuti per ciascuna prova) la prova viene considerata
irrisolta, dal momento che il criterio stabilito per il compito non
è stato raggiunto. L'Analisi della Varianza per Trend è stata condotta
per i tempi di "Latenza di attacco", di "Operandum", e di " Detour"
sull'intero gruppo di animali.
Considerando
l'intero campione si evidenza un F altamente significativo ( P<0.001)
sia nell'Operandum" che nel "Detour" (Operandum: F=2.342 con 19 e
361 gl). Per i tempi di "Latenza di attacco" non risulta alcuna differenza
significativa tra le prove (F= 1.396 con 19 e 361 gl).
Considerando separatamente i tre sottogruppi di animali, appare chiaro come la significativtà dei valori di F, relativi ai tempi di "Operandum" e "Detour", riscontrata sull'intero campione di animali, sia determinata per la maggior parte dalla forte significatività del trend nel sottogruppo appartenente alla terza fase sperimentale (Operandum gruppo 1:F=0.575 con 19 e 114 gl; Detour gruppo 1: F= 0.448 con 19 e 114 gl; Operandum gruppo 2: F= 1.256 con 19 e 76 gl; Detour gruppo 2:F= 0.898 con 19 e 76 gl. Operandum gruppo 3: F= 3.489 con 19 e 133 gl; Detour gruppo 3: F= 3.148 con 19 e 133 gl).
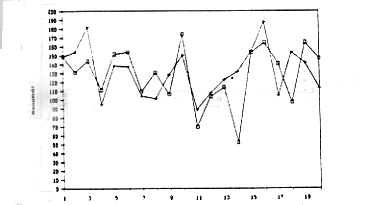
Figura 4
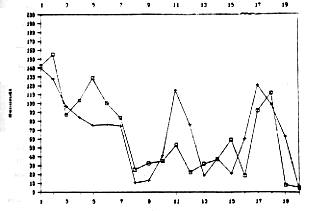
Figure
4 e 5 - Andamento delle medie dei tempi di Operandum (contrassegnati
con un più) e Detour (contrassegnati con un quadratino) rispettivamente
per gli animali del primo e del secondo gruppo.
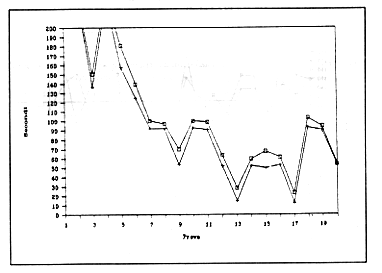
Figura
6 - Andamento delle medie dei tempi di Operandum (contrassegnati
con un più) e Detour (contrassegnati con un quadratino) per
gli animali del terzo gruppo.
Sono anche state considerate eventuali differenze relative alle variabili peso e sesso nell'analisi della prestazione globale. I risultati non evidenziano alcuna significatività della relazione Prestazione-Peso, probabilmente a causa della insufficiente distribuzione tra le classi di peso dei soggetti (Fig. 7 e 8).
Per quanto riguarda, poi la relazione Prestazione-Sesso, dai dati non risulta una differenza significativa (test di Mann-U-Withney: N.S.) tra i due sessi.
Debita attenzione deve essere dedicata alle prestazioni dei singoli soggetti.
Ovviamente,
nella valutazione è necessario tener conto del fatto che Octopus
vulgaris è un invertebrato, cioè un animale con elevata variabilità
comportamentale e quindi "psicologica".
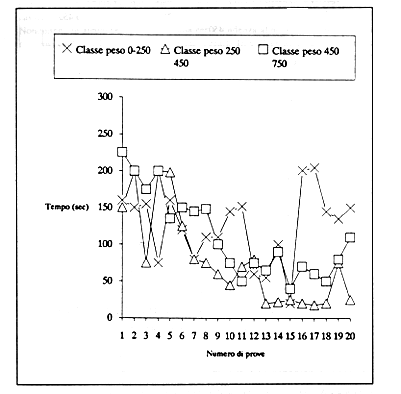
Figura
7 - Andamento delle medie dei tempi di Operandum degli
animali suddivisi in tre classi di peso. I grafici non dimostrano
una significativa differenza negli andamenti tra le classi di peso
analizzate.
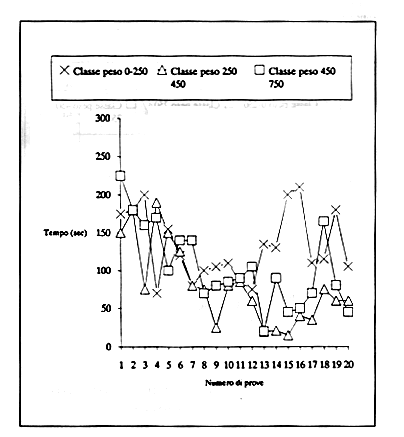
Figura 8 - Andamento delle medie dei tempi di Detour degli animali suddivisi in tre classi di peso . I grafici non dimostrano una significativa differenza negli andamenti tra le classi di peso analizzate.
La Figura 9 riporta le prestazioni dei singoli soggetti utilizzati nella sperimentazione. Risulta evidente come esistano, anche all'interno di ciascun gruppo, delle forti differenze tra gli individui. All'interno del primo gruppo (Fase 1) vi sono animali che non hanno mai, o quasi, risolto il problema (Animale n.11), o che dall'andamento del grafico sembrano aver risolto il compito in maniera del tutto casuale (Animali n.10 e 14).
Nondimeno in alcuni Octopus risulta una certa tendenza al miglioramento, anche se non così evidente: l'animale n. 9 nel Detour tra la II e III prova e nell'Operandum tra l'VIII e la XIII prova; il n. 12 nel Detour tra la V e la IX prova dell'Operandum tra la VI e l'VIII; il n. 13 nel Detour tra la III e la VI prova nell'Operandum tra la IV e la VII prova.
Nel secondo
gruppo (Fase 2) particolarmente l'animale n. 16 ed il n. 21 mostrano
un miglioramento progressivo della prestazione nell'Operandum e nel
Detour durante la prima metà dell'esperimento. Nel terzo gruppo (Fase
3), infine, le curve dimostrano con maggior evidenza l'apprendimento,
pur restando una notevole diversità tra le prestazioni degli individui.
Ad es, l'animale n. 35 non ha mai risolto nelle prime 12 prove (a
causa di inattività, come risulta evidente dall'assenza di attacchi);
a partire dalla XIII prova mostra un progressivo miglioramento nel
compito di Detour. Anche negli animali n.32, 37 e38 nelle fasi centrali
dell'esperimento c'è una particolare evidenza di miglioramento preogressivo
nel compito di Detour. L'animale n.29 a partire dalla V prova mostra
una chiarissima tendenza sia nel Detour che nell'Operandum. Anche
l'Octopus n. 34 nella seconda metà dell'esperimento migliora
progressivamente in entrambe le fasi del problema, mentre il n. 36
migliora particolarmente nell'Operandum.
Dai risultati sopra indicati, ed in particolare da quelli relativi al terzo gruppo, risulta evidente la capacità di apprendimento in Octopus vulgaris di entrambe le fasi di cui si compone il compito.
Considerando
l'intero campione di soggetti (n.=20), il numero di prove irrisolte
diminuisce gradualmente con una punta minima alla XIII prova, nella
quale solo 2 animali su 20 non risolvono il compito. Il numero di
prove irrisolte può essere considerato come in indice generale di
insuccesso, che però non dice nulla circa le cause per cui un certo
numero di prove non vengono risolte. Potrebbe trattarsi tanto dell'inattività
di alcuni animali, quanto di una attività caratterizzata da un elevato
numero di errori. Ci si riferisce, dunque, al totale dei tentativi
effettuati da tutti gli animali in ciascuna prova prima di risolverla.
Tale percentuale si riduce progressivamente dal 60-65% di attacchi
falliti sul totale nelle prime tre prove, fino a raggiungere la sua
punta minima, inferiore al 20%, nella settima ed ottava prova.
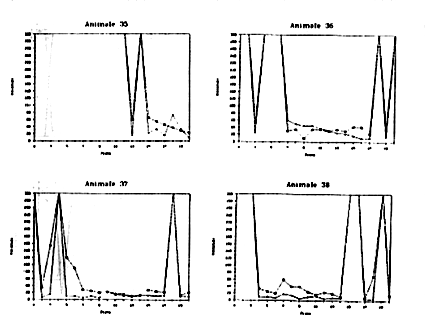
Figura 9a
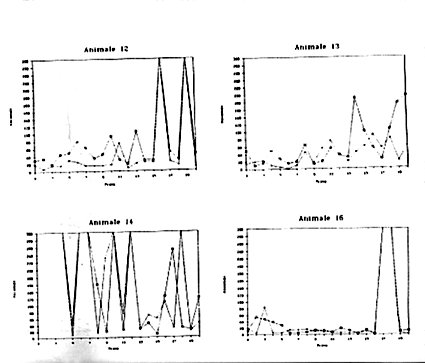
Figura 9b
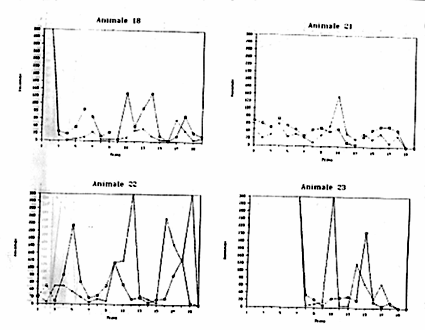
Figura 9c
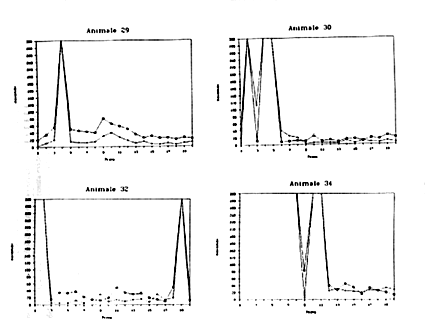
Figura 9d
Figura 9e
Figure9a,
9b, 9c, 9d, 9e, -Andamento delle prestazioni di Operandum
(indicato con una crocetta) e di Detour ( indicato con un quadratino)
per alcuni esemplari di Octopus vulgaris.
Oltre ad una riduzione degli attacchi falliti, si assiste anche ad un altro tipo di miglioramento: nel corso delle prime prove, gli Octopus riducono progressivamente il numero di errori, effettuando meno tentativi prima di riuscire a risolvere il compito, ed i loro comportamenti di attacco tendono più all'azione sul cilindro che non ad afferrare il granchio in esso contenuto.
Quest'ultima affermazione risulta evidente anche da una preliminare analisi del materiale videoregistrato sulle strategie adottate dagli Octopus.
Solitamente,
nel corso delle prime prove, gli animali attaccano il barattolo dapprima
avvicinandosi con cautela spostandosi lungo la vasca, o a una lato
di essa, per azione dei tentacoli. Giunti ad una distanza di 20-30
cm gli Octopus, con "balzo" a tentacoli semiaperti, tentano
la presa del granchio ritirandosi immediatamente di fronte ad un ostacolo
imprevisto ed invisibile. In molti animali si assiste, a questo punto,
ad un iniziale incremento del numero di attacchi e ritirate, eseguiti
in rapida successione, ma sempre mirati alla preda, e talvolta caratterizzati
da una certa attività esplorativa con uno o due tentacoli. Inizialmente
a caso, alcuni animali risolvono il compito esplorando il barattolo
dall'alto ed emettendo una risposta consumatoria nei confronti del
tappo, che in tal modo viene sollevato. Altri polpi capovolgono il
barattolo, spesso trascinandolo verso la tana, esplorandolo da tutti
i lati fino a tirarne casualmente il coperchio. L'attività esplorativa
si riduce sempre più. Accade spesso che gli animali mantengano anche
successivamente, nella maggioranza delle prove, lo stesso schema comportamentale
con il quale hanno ottenuto la preda la prima volta (ad esempio capovolgendo
sempre o non capovolgendo mai il barattolo), ma in ogni caso cessano
progressivamente di dirigere i propri attacchi alla preda, attaccando
direttamente il tappo o allungando direttamente alcuni tentacoli verso
la parte superiore del cilindro per tirarlo e capovolgerlo.
A questo
punto è necessario domandarsi fino a che punto sia lecito parlare
di una associazione tra la sequenza comportamentale che immediatamente
precede il successo, come suggerirebbe il fatto che la medesima strategia
tende ad essere mantenuta nelle varie prove. Un'interpretazione unicamente
in termini di associazione S-R appare insufficiente: certamente alla
base di questo tipo di apprendimento (specialmente nel "Detour") vi
è l'acquisizione delle caratteristiche del barattolo, e sarebbe interessante
verificare l'eventuale effetto di un preallenamento esplorativo sulla
rapidità del processo di apprendimento.
Circa le differenze tra le prestazioni dei tre diversi schemi sperimentali, si può senz'altro concludere che la prossimità temporale tra le prove è un elemento importante se non addirittura determinante. Infatti confrontando le Figure 4 e 5 con la Figura 6 ci si rende conto come l'alta significatività del trend nel terzo gruppo sia imputabile al fatto che le sessioni giornaliere erano due e non una sola (come nel secondo gruppo). Queste differenze tra le tre fasi sperimentali sono imputabili alle diverse condizioni dell'esperimento.
La non significatività nella prima fase è da imputare ad una caduta della prestazione riscontrabile verso le ultime prove nella maggior parte degli animali (in particolare n.8,9,12 e 13). La caduta motivazionale è imputabile probabilmente all'incidenza della temperatura che, essendo particolarmente bassa (inverno pieno: cfr. Tabella 3), rallenterebbe il metabolismo degli Octopus, riducendone la necessità di cibo.
La non significatività del trend nel secondo sottogruppo (in cui a motivo dei risultati della prima fase era stato abbassato a uno il numero delle sessioni giornaliere), appare determinato sia da una caduta nelle prestazioni finali in alcuni animali ( ad es. il n.16), sia ad una disomogeneità delle prestazioni nella maggioranza dei soggetti. Evidentemente la maggiore distanza in termini di tempo da una prova all'altra deve avere avuto un suo peso nel processo di ritenzione dell'esperienza precedente. Il dato della forte significatività del trend nel terzo sottogruppo è chiaramente risultato di una condizione sperimentale favorevole. La grande differenza dovuta all'effetto della variabile temperatura risulta chiara confrontando ancora le Figure 4 e 6.
Riguardo alla variabile "sesso", dai testi effettuati non risulta alcuna differenza significativa tra i maschi e le femmine del campione così pure la variabile "peso", nell'intervallo considerato non sembra costituire un fattore che incide sulla prestazione, ma piuttosto può incidere sull'adattamento dell'animale alla vita in acquario. I soggetti di taglia maggiore sono quelli che maggiormente presentano difficoltà in tal senso: ad esempio gli animali 14 (570 g), 32 (690 g) e 35 (750 g) tendevano ad essere più inattivi nelle prime prove dell'esperimento
In conclusione si può affermare che l'Octopus vulgaris è in grado di migliorare il proprio adattamento ambientale, apprendendo la soluzione di un compito complesso, quale è quello rappresentato da un susseguirsi di azioni che vanno dall'attacco ad un contenitore, fino all'aggiramento di un ostacolo costituito dal tappo del contenitore che deve essere tolto per dare accesso alla cattura della preda e quindi all'atto consumatorio del nutrirsi che chiude ed esaurisce la fase motivazionale. Inoltre intervengono fatti mnemonici, sempre connesso al fenomeno dell'apprendimento, come mostrano gli andamenti del secondo gruppo comparati a quelli del terzo ( Figure 5 e 6 ), nel quale l'Octopus ha la possibilità di produrre risultati su un'insieme di prove ravvicinate. I polpi sono in grado di ritenere quegli schemi a catene comportamentali che, nel presente esperimento, rappresentano una soluzione adattiva alle condizioni imposte dall'ambiente. Probabilmente tale compito ( afferrare il vasetto di vetro, togliere il tappo e inserire il tentacolo per prendere la preda) costituisce una prova relativamente facile per una specie dotata di braccia, particolarmente atte alla manipolazione. Ciò non di meno gli andamenti dei risultati mostrano che tali condotte non sono dovuti all'azione di moduli stereotipati e ciechi, ma all'uso "cognitivo" ed alaborato delle capacità motorie. In altre specie, soprattutto se tassonomicamente superiori, in particolare nei Primati e nell'uomo, noi siamo abitualmente condizionati a considerare simili condotte come "intelligenti".
Nel continuum della scala evolutiva Octopus sembra essere uno dei primi gradini della manifestazione di fenomenologie comportamentali intelligenti. Non v'è da stupirsi troppo, poiché ciò che etichettiamo col termine intelligenza, come ciò che etichettiamo con i termini di apprendimento o memoria, è una proprietà della materie vivente organizzata in sistemi complessi. Apprendimento e memoria sono proprietà della materia organica già a livello di organizzazione di macromolecole. Gli organismi pluricellulari sono tutti sistemi complessi, se si considera che già una singola cellula è un sistema dotato di diverse e differenziate strutture in grado di svolgere molteplici funzioni. Quando poi l'organismo ha un sistema nervoso consistemente sviluppato, (ed è poi il caso dell'Octopus) il livello comportamentale dello psichismo e le relative funzioni sono sicuramente raggiunti. Sembra infatti, sempre più chiaramente col progredire delle ricerche l'estendersi delle conoscenze (soprattutto comparative), che le differenze maggiori siano più a livello delle quantità che delle qualità. Le medie della diminuzione progressiva del numero di prove irrisolte e la riduzione degli attacchi fallimentari insieme al progressivo ridimensionamento degli errori, costituito da un minor numero di tentativi prima di risolvere il compito, ma anche della direzione dell'azione sempre più rivolta ad operare sul vasetto di vetro invece che nel tentare di prendere il granchio attraverso il vetro, indicano chiaramente il progredire, attraverso l'apprendimento, della comprensione "intelligente" e non casuale del problema e della strategia migliore per risolverlo.
L'apparente disparità dei dati presentati in questa ricerca rispetto alla letteratura esistente, pur confermando la capacità di Octopus vulgaris di apprendere, stabilisce un nuovo modello sperimentale teso allo studio dell'abilità generale della specie, della variabilità individuale e dei processi attraversi i quali questa specie traduce le proprie potenzialità adattive, modificando stabilmente il proprio comportamento. Il fatto che gli studi di Piéron (1911), Boycott (1953) e Wells (1964) sull'apprendimento di Octopus, indipendentemente delle tecniche sperimentali, abbiamo tenuto conto esclusivamente delle prestazioni dei "migliori" soggetti, ha probabilmente "nascosto " agli occhi di questi ricercatori l'evidenza della diversa adattabilità individuale ella condizioni sperimentali e probabilmente a quelle naturali. Lo studio più analitico delle prestazioni comportamentali e delle strategie adottate in ciascuna risoluzione potrà certamente dare maggiori indicazioni circa le capacità oggettive di Octopus vulgaris a risolvere quindi compiti complessi e sul ruolo che l'apprendimento può svolgere nel suo habitat naturale.

